Overview
These two layers of cell growth are interconnected, with signaling pathways controlling metabolism, and metabolites controlling signaling.
The Teleman Lab is interested in understanding cell and tissue growth. Compared to the cell cycle, the mechanisms controlling cell growth and tissue growth are less well understood.
We study two aspects: metabolism and signaling.
Metabolism: To grow, cells and tissues need to make biosynthetic building blocks to accumulate mass. This requires the activation of metabolic pathways that support anabolism and fight oxidative stress.
Signaling: Cell metabolism, in turn, is regulated by signaling pathways such as the insulin pathway or the mTORC1 pathway. These signaling pathways sense and respond to growth factors and nutrient levels. We investigate the regulation of these signaling processes controlling metabolism and energy consumption.
Our work has been supported by two ERC grants.
This research has implications for both basic biology and cancer biology:
Basic biology: Tissue growth and size control represents one of the unsolved fundamental mysteries in developmental biology. How does a tissue know what size to become, and when to stop growing? How does a tissue measure its size? The underlying mechanisms are not known.
Cancer biology. To form a tumor, cells need to both proliferate and grow. Understanding the molecular mechanism controlling cell growth, and the metabolic pathways that generate biomass, is an important part of understanding cancer.
Model systems. Since tissue growth control is a conserved process in all animals, we use both Drosophila and human cells in culture as model systems. We use Drosophila mainly for gene discovery and for organismal studies. We use mammalian cell culture to study basic biological mechanisms, to screen for genes involved in processes of interest, and to translate our findings from flies into mammals. We also study patient samples with our collaborators at the Heidelberg University Hospital.
Neurofibromatosis Type II and Schwannomas
Finding a therapy for NFII
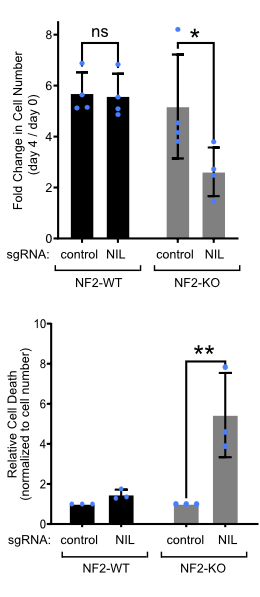
Synthetic lethality of NF2 and NIL
© dkfz.de
Neurofibromatosis Type II (NFII) is a genetic condition characterized by predisposition to benign tumors of the peripheral nervous system. It is caused by homozygous loss-of-function of the homonymous gene NF2 in Schwann cells, the glia that ensheath peripheral nerves. Individuals with NFII typically harbor a germline mutation in one of the two NF2 alleles. Recurrent, stochastic, somatic loss of the second NF2 allele gives rise to tumors throughout the lifetime of the patients. These tumors grow and constrict the nerves. Since cranial nerves are often affected, this leads to balance problems, dizziness, headaches, facial weakness, and loss of hearing or sight. Unfortunately, there is no approved pharmacological therapy for NFII. Tumors are surgically resected, leading to nerve damage. Since patients recurrently develop new tumors, they need to return to the hospital for tumor resections every few years, leading to significant morbidity.
We are working on developing a pharmacological therapy for NFII. To this end, we performed a screen to search for genes that, when knocked-down or inhibited, cause death of NF2 mutant Schwann cells (ie the tumor) but not of NF2 wildtype or NF2 heterozygous Schwann cells (ie the rest of the patient). This identified a target which we call NF2-Interacting-Lethal partner (NIL). Inhibition of NIL induces death of NF2 knockout but not NF2 wildtype Schwann cells (see figure to the right). This means that if we develop a drug that inhibits NIL, we should be able to kill NF2 mutant tumors. Indeed, in work done together with the DKFZ xenograft core facility, we found that NIL inhibition kills NF2 mutant xenografts in mice.
We are now developing a drug targeting NIL together with Evotec and Bristol Myers Squibb in a beLAB2122 project.
Discovering new genes that cause Schwannomas
As a side-product of the screen we did for genes regulating Schwann cells proliferation and survival, we also discovered several new genes that act as tumor suppressors in Schwann cells. We are now working on them to characterize their phenotype, to understand their molecular mode of action, and to understand whether they are mutated in human Schwannomas. We hope this work will uncover genes that are responsible for the initiation of Schwannomas, improving our understanding of this disease, and identifying new approaches for targeting the disease.
Diabetes
Reactive carbonyl species and causes of diabetes
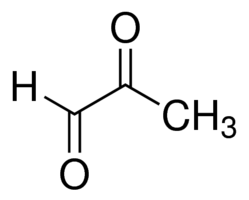
Methylglyoxal (MG)
Type 2 diabetes is associated with insulin resistance and hyperglycemia so much so, that insulin resistance is often considered to be the cause of type 2 diabetes. Indeed, Type 2 diabetes is treated with insulin sensitizers or with insulin. Unexpectedly, however, recent clinical studies find that therapies renormalizing blood glucose to non-diabetic levels does not prevent progression of the disease patients still develop neuropathies, nephropathy and retinopathy. This suggests that insulin resistance is one phenotype of type 2 diabetes, but not the cause. This raises the important question: what are the causes of type 2 diabetes?
We are working on this question in collaboration with the Szendrödi lab at Heidelberg University Hospital. Data from our lab suggest one possible cause could be elevated levels of the metabolite methylglyoxal. Methylglyoxal, which forms as a byproduct of several metabolic pathways, forms adducts on proteins to alter their function. We are studying the detoxification of methylglyoxal, and the consequences of impaired detoxification for diabetes.
Selected reference
Moraru A, Wiederstein J, Pfaff D, Fleming T, Miller AK, Nawroth P and Teleman AA. Elevated levels of the reactive metabolite methylglyoxal recapitulate progression of Type 2 Diabetes. (2018) Cell Metabolism doi: 10.1016/j.cmet.2018.02.003 [pubmed]
Pfaff DH, Fleming T, Nawroth P and Teleman AA. Evidence against a role for the Parkinsonism-associated protein DJ-1 in methylglyoxal detoxification. (2016) JBC 292:685-690 [pubmed]
Basic / Fundamental Biology
Understanding growth control during development
Wing size is tightly controlled in Drosophila.
A fundamental question in developmental biology is to understand how organs can measure and regulate their size. For instance, the two legs of a person or the two wings of a fly are usually identical in size to within 1%. How this is controlled at the molecular level is a mystery.
During animal development tissues increase tremendously in mass, growing roughly 1000-fold from the embryo to the adult. Each tissue then stops at a very precise size, which is determined by a combination of genetic and environmental factors. We try to unravel the molecular machinery controlling this termination of growth. We believe this is relevant for cancer biology, where tumors do not respect the regulation of correct organ size.
The best studied model system for growth control is the fly wing due to the extensive genetic tools available to manipulate gene expression in time and space. We use this system to study how signaling pathways control growth, such as the (1) Insulin / PI3K / mTOR, (2) Hippo / Yorkie/YAP, and (3) Dpp (TGF-beta) pathways.
Selected references
Strassburger K, Lutz M, Müller S and Teleman AA. Ecdysone regulates Drosophila wing disc size via a TORC1 dependent mechanism. (2021) Nature Communications. 12(1):6684 [pubmed]
Romero-Pozuelo J, Demetriades C, Schroeder P and Teleman AA. CycD/Cdk4 and discontinuities in Dpp signaling activate TORC1 in the Drosophila wing disc. (2017) Developmental Cell 42:376-87 [pubmed]
Doumpas N, Ruiz-Romero M, Blanco E, Edgar B, Corominas M and Teleman AA. Brinker regulates wing disc growth in part via repression of Myc expression. (2013) EMBO Reports. 14:261-8 [pubmed]
Teleman AA, Cohen SM, Dpp gradient formation in the Drosophila wing imaginal disc. (2000) Cell. 2000 Dec 8;103(6):971-80 [pubmed]
Dynamic regulation of TOR by amino acids
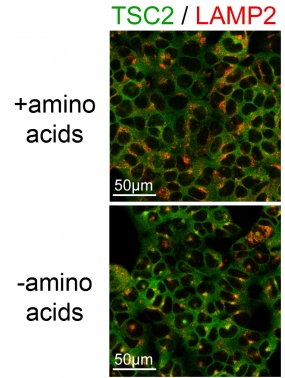
The images show the TSC2 localization in cells by immunostaining of TSC2 (green) and the lysosomal marker LAMP2 (red). Amino acid starvation (-amino acids) causes lysosomal re-localization of TSC2 (yellow).
© dkfz.de
mTORC1 is a central regulator of cell growth. Highly conserved from yeast to humans, it is one of the most powerful anabolic signals in a cell. When mTORC1 is active, cells accumulate mass and grow, whereas when mTORC1 is inactive, cells cannot grow.
For this reason, mTORC1 is dysregulated in almost all human cancers. mTORC1 promotes cell growth by inducing protein synthesis, as well as synthesis of other cellular building blocks such as lipids for the cell membrane. Because of its central role in regulating growth, mTOR activity is regulated by a large number of inputs including growth factor signaling (such as insulin), cellular stresses (such as hypoxia, or high AMP levels) as well as the availability of amino acids.
The molecular mechanisms by which some of these inputs regulate mTOR, however, are still not understood. We found that both amino acids and growth factors are required to avoid mTORC1 inhibition by the TSC1/2 complex. The presence of any single stress is sufficient to cause TSC2 lysosomal localization and inhibition of TORC1. This effect is not cell-type specific, indicating an underlying general biological detection mechanism to a variety of stress stimuli. We also found a mechanism that spares neurons from inhibiting mTORC1 which nutrients are low. This is important so that our brain neurons don't become catabolic if we skip a meal !
A few projects in the lab aim at identifying novel regulators of TOR, in particular relating to cellular amino acid sensing, and to understand their function both at the molecular and the physiological level.
Selected references
Demetriades C, Doumpas N, and Teleman AA. Regulation of TORC1 in Response to Amino Acid Starvation via Lysosomal Recruitment of TSC2. (2014) Cell. 156:786-99 [pubmed]
Plescher M, Teleman AA and Demetriades C. TSC2 mediates hyperosmotic stress-induced inactivation of mTORC1. (2015) Scientific Reports 5:13828 [pubmed]
Tsokanos F-F, Albert M-A, Demetriades C, Spirohn K, Boutros M, and Teleman AA. eIF4A inactivates TORC1 in response to amino acid starvation. (2016) EMBO J. 2016 May 17;35(10):1058-76 [pubmed]
Demetriades C, Plescher M, and Teleman AA. Lysosomal recruitment of TSC2 is a universal response to cellular stress. (2016) Nature Communications 7:10662. [pubmed]
Figlia G, Müller S, Hagenston AM, Kleber S, Roiuk M, Quast JP, Ten Bosch N, Carvajal Ibañez D, Mauceri D, Martin-Villalba A, and Teleman AA. Brain-enriched RagB isoforms regulate the dynamics of mTORC1 activity through GATOR1 inhibition. (2022) Nature Cell Biology. [pubmed]
Cancer and non-canonical mRNA translation

DENR-KO die as pharate adults with larval-like abdominal epidermis. DENR promotes cell proliferation and boosts protein synthesis in proliferating cells.
© dkfz.de
To grow, cancer cells need to synthesize proteins. Cancer cells, however, face a challenge. Canonical textbook cap-dependent translation is turned off in response to cellular stresses. Hypoxia, nutrient limitation, ER stress, genotoxic stress and cytotoxic stress all inhibit canonical translation (by phosphorylating eIF2alpha and by inhibiting mTORC1, which leads to 4E-BP activation and eIF4E inhibition). Since almost all cancer cells have one stress or the other, they need to either bypass these inhibitory mechanisms, or to rely on alternate non-canonical translation mechanisms. We are studying some of these non-canonical translation mechanisms.
For instance, we discovered that the oncogenes DENR and MCTS1 promote a process called translation re-initiation, to promote expression of genes required for proliferation. Several projects in the lab focus on how translation is regulated, and how its dysregulation contributes to cancer.
Selected references
Ahmed YL, Schleich S, Bohlen J, Mandel N, Simon B, Sinning I, Teleman AA. DENR-MCTS1 heterodimerization and tRNA recruitement are required for translation reinitiaion. (2018) PLoS Biology 16(6):e2005160. [pubmed]
Bohlen J, Harbrecht L, Blanco S, Clemm von Hohenberg K, Fenzl K, Kramer G, Bukau B and Teleman AA. DENR promotes translation reinitiation via ribosome recycling to drive expression of oncogenes including ATF4. (2020) Nature Communications 11:4676. [pubmed]
Bohlen J, Fenzl K, Kramer G, Bukau B and Teleman AA. Selective 40S footprinting reveals cap-tethered ribosome scanning in human cells. (2020) Molecular Cell 79:561-574.e5 [pubmed]
Schleich S, Strassburger K, Janiesch PC, Koledachkina T, Miller KK, Haneke K, Cheng Y-C, Kuechler K, Stoecklin G, Duncan KE and Teleman AA. DENRMCT-1 Promotes Translation Reinitiation Downstream of uORFs to Control Tissue Growth. (2014) Nature 512: 208-212 [pubmed]
Schleich S, Acevedo JM, Clemm von Hohenberg K and Teleman AA. Identification of transcripts with short stuORFs as targets for DENRMCTS1-dependent translation in human cells. (2017) Scientific Reports 7:3722 [pubmed]